Modifications of Aortic Arches in Vertebrates
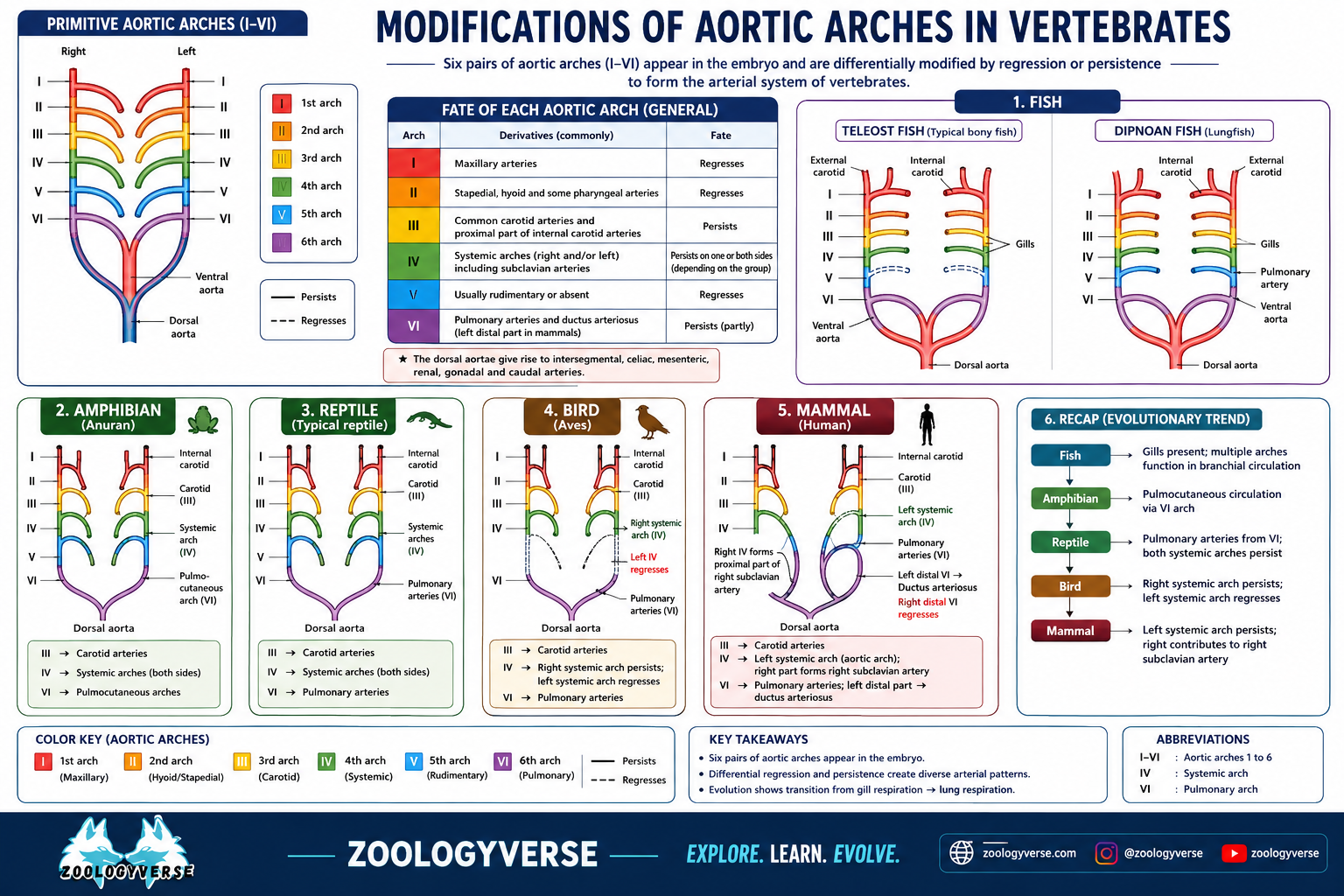
Modifications of Aortic Arches in Vertebrates :- Basic embryonic plan. In a typical vertebrates embryo, the major arterial channels include a ventral aorta, a dorsal aorta and usually 6 pairs of aortic arches connecting ventral aorta with the dorsal aorta. Blood leaves the heart through ventral aorta which runs forward, midventrally beneath the pharynx and branches anteriorly into a pair of external carotid arteries into head. Ventral aorta gives off, at intervals, 6 pairs of aortic arches running through the visceral arches. Each aortic arch consists of a ventral afferent branchial artery carrying venous blood to capillaries in a gill, and a dorsal efferent branchial artery taking arterial blood from the gill. All the efferent branchial arteries of the same side dorsally join a lateral dorsal aorta or radix which is extended into head as the internal carotid artery. The two lateral dorsal aortae unite just behind the pharynx to form a single median dorsal aorta which continues behind into tail region as caudal artery. Branches from these main arterial channels supply all parts of the vertebrate body.
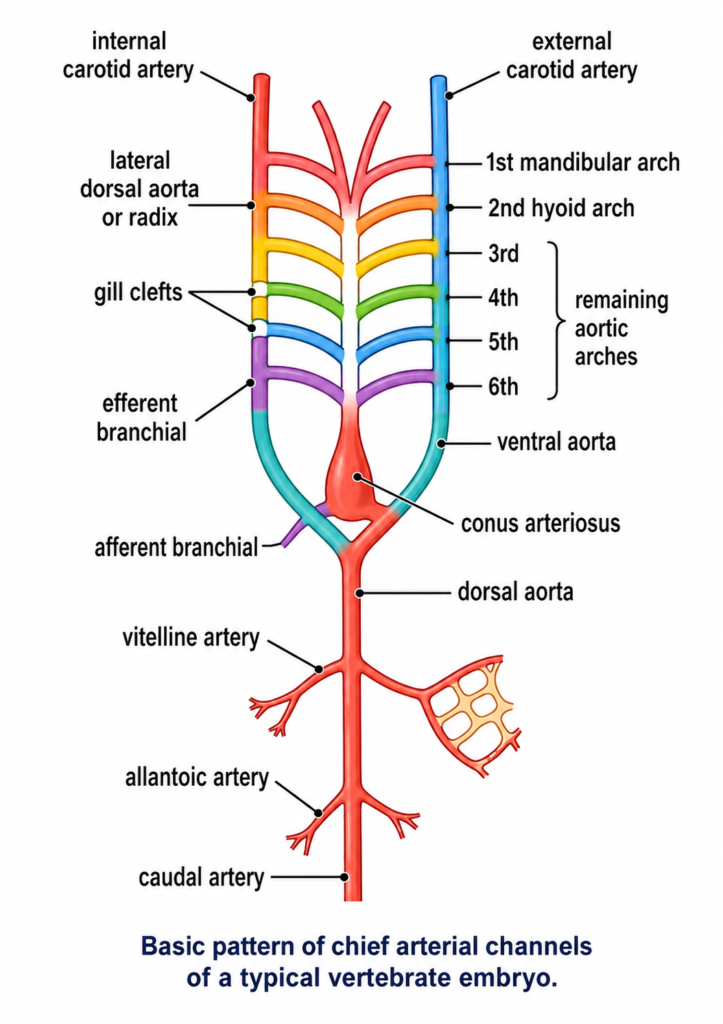
Although arterial system of different adult vertebrates shows major differences, but it is actually built according to the same basic architectural plan as seen in the vertebrate embryo. The differences are due to increasing complexity of heart on account of a shift from gill respiration to lung respiration. The modifications mainly concern the aortic arches which undergo a progressive reduction in number from lower to higher vertebrates.
Primitive vertebrates :- In Branchiostoma (amphioxus), nearly 60 pairs of aortic arches are present, connecting the ventral and dorsal aortae. In Petromyzon, 7 pairs of aortic arches are found. In other cyclostomes the number varies from 6 (Myxine) to 15 pairs (Eptatretus).
Fishes :- The primitive elasmobranch (Heptanchus) has 7 pairs of aortic arches. Most of the fish embryos present primitive plan with 6 or more pairs of aortic arches, each passing through a gill. But, in adult condition, the number is reduced to 4 or 5. In most sharks (elasmobranchs), only 5 pairs (II, III, IV, V, and VI) are functional. The first gill slit forms the spiracle which is non-functional as a gill. Accordingly the first arch (mandibular) is absent or represented by an efferent pseudobranchial artery. In most teleosts or bony fishes, I and II arches tend to disappear, so that only 4 pairs (III, IV, V and VI) remain functional. In Polypterus and lungfishes (Dipnoi), gills are poorly developed, so that a pulmonary artery arises from the efferent part of the VI arch on each side and supplies blood to the developing air bladder or lung. In Protopterus, the III and IV embryonic arches are uninterrupted by gill capillaries.
In elasmobranchs and lungfishes, each arch forms one afferent and two efferent arteries (by splitting) in each gill. In teleosts or bony fishes, each gill has one afferent and one efferent artery.
In tetrapods, true internal gills are absent so that aortic arches do not break up into afferent and efferent arteries. I and II arches totally disappear in all tetrapods.
Amphibians :- With the introduction of lungs as main respiratory organs and the diminishing importance of gills, the aortic arches of amphibians show a modification from those of fishes. Urodeles or the tailed amphibians live in water and retain external gills permanently in addition to lungs. Accordingly, their aortic system shows only partial shift from condition in fishes. 4 pairs of arches (III to VI) are usually present, although in some forms (Necturus, Siren, Amphiuma), V arch is incomplete, reduced or absent. Thus tailed amphibians show transition from 4 to 3 pairs of aortic arches. Ill arch forms the carotid arches, IV the systemic arches. The radix or lateral aorta between III & IV arches may persist as a vascular connection termed ductus caroticus. VI arch on either side becomes the pulmocutaneous artery or arch, supplying blood to skin and lungs. However, it also retains connection with radix aorta called ductus Botaili or ductus arteriosus. In the larval stage of an anuran or tailless amphibian, such as frog tadpole, arrangement of aortic arches is similar to an adult urodele, due to gill respiration. At metamorphosis, with loss of gills, aortic arches I, II and V disappear altogether. Ductus caroticus also disappears so that the III or carotid arch takes oxygenated blood only to head region. IV or systemic arch on each side continues to dorsal aorta to distribute blood elsewhere except head and lungs. Ductus arteriosus also disappears so that VI or pulmocutaneous arch supplies venous blood exclusively to lungs and skin for purification. Thus, adult anurans exhibit only 3 functional arches, (III, IV and VI) which are also retained by the amniotes or higher vertebrates.
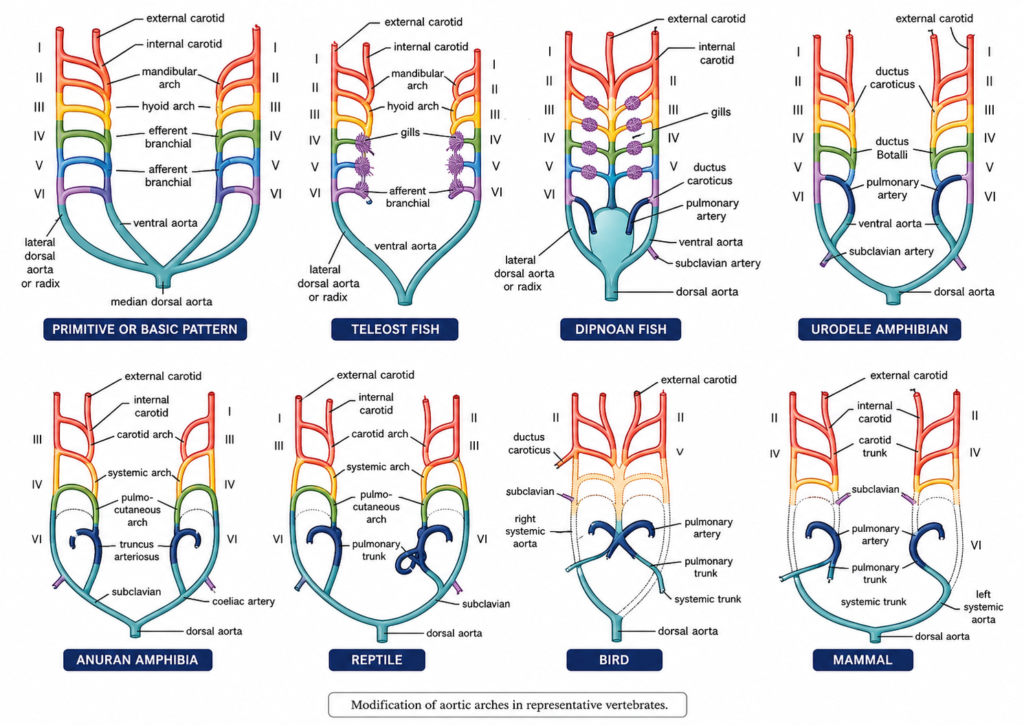
Reptiles :- Reptiles vertebrates in are fully terrestrial which gills disappear altogether and replaced by lungs. Only 3 functional arches (in, IV and VI) are present. But elongation of neck, posterior shifting of heart and partial division of ventricle brings about certain innovations in the aortic system.
- Entire ventral aorta and conus split forming only 3 trunks-two aortic or systemic and one pulmonary.
- Right systemic arch (IV) arises from left ventricle carrying oxygenated blood to the carotid arch (HI) to be sent into head.
- Left systemic arch (IV) leads from right ventricle carrying deoxygenated or mixed blood to the body through dorsal aorta.
- Pulmonary trunk (VI) also emerges from right ventricle carrying deoxygenated blood to the lungs for purification.
- Ductus caroticus and ductus arteriosus are absent. But, ductus caroticus is present in certain snakes and lizards (Uromasitx), ductus arteriosus in some turtles, and both in Sphenodon. Reptiles also remain cold-blooded, like amphibians and fishes, due to mixing of blood.
Birds and mammals :- Birds and mammals are warm-blooded because in both the ventricle is completely divided so that there is no mixing of oxygenated and unoxygenated bloods. As usual, 6 arches develop in the embryo, but only 3 arches (III, IV, VI) persist in the adult. Other features are as follows —
- Ventral aorta is replaced by two independent aortae or trunks-systemic and pulmonary.
- Arch IV is represented by a single systemic aorta, right in birds and left in mammals, emerging from left ventricle and carrying oxygenated blood. Uniting with the radix aorta of its side it forms the dorsal aorta.
- The only remaining part of the other lost systemic arch is represented by a subclavian artery, on left side in birds and on right side in mammals.
- Arch III with remnants of lateral and ventral aortae represents carotid arteries, which arise from systemic aorta.
- Arch VI forms a single pulmonary taking deoxygenated blood ventricle to the lungs.
- Embryonic ductus caroticus from and trunk right ductus arteriosus also disappear. The latter closes but persists until hatching or birth in some cases as a thin ligament of Botalli or ligamentum arteriosum.
Venous System
Deoxygenated or venous blood from different parts of the body is returned to the heart via veins. Like arteries, the veins of all vertebrates also follow a basic pattern or fundamental plan.
[I] Embryonic veins
The venous system in early embryonic life of all vertebrates is relatively simple, similar and in accordance with the basic pattern. Most of the veins are paired and symmetrically arranged. The major basic embryonic veins include : Cardinals (anterior, posterior, and common cardinal or ductus Cuvieri), lateral abdominal, subintestinal, and caudal.
[II] Modifications of veins in vertebrates
Modifications in adult vertebrates occur by either deletions or additions of some veins to the basic embryonic pattern Modifications are few in elasmobranchs but more numerous in tetrapods. In vertebrates, veins can be arranged in three distinct categories- systemic or portal somatic, renal and hepatic portal. A fourth category of pulmonary veins and postcaval veins in added in lungfishes and tetrapods.
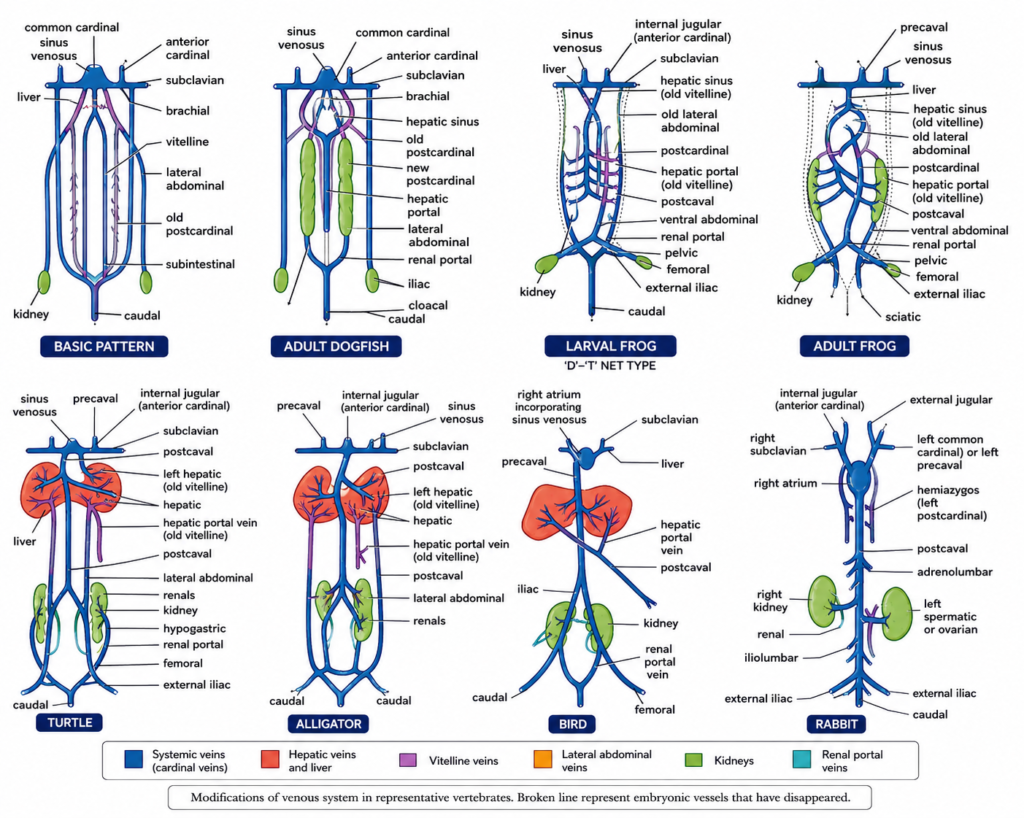
1. Systemic veins. Systemic or somatic veins collect blood from all parts of the body and empty into sinus venosus of the heart.
- Elasmobranchs :- In adult cartilaginous fishes (dogfish), venous system is almost a blueprint of the basic architectural plan of the embryo. A few larger veins expand to form thin-walled sinuses. Blood from head and posterior region of body is collected by large, paired anterior and posterior cardinal veins respectively. On either side they open into the common cardinal or ductus Cuvieri, that passes inwards through transverse septum to enter the sinus venosus of heart. In fishes and salamanders, an inferior jugular vein also collects blood on either side from ventral part of head to join the common cardinal. In embryo, posteriorly, the two posterior cardinal veins remain continuous with a caudal vein collecting blood from tail. Each posterior cardinal, or postcardinal, runs anteriorly along the outer margin of kidney, draining it through a series of renal veins, before joining the common cardinal. In adult dogfish, the old postcardinals become interrupted near anterior ends of kidneys. Instead new postcardinals (earlier subcardinal channels) develop along the inner margins of kidneys which they drain. Blood from lateral wall and pelvic fin on either side is returned through a ventral or lateral abdominal vein. It receives a branchial vein from pectoral fin forming a short subclavian vein enters the common cardinal of its side. Abdominal veins are absent in bony fishes or teleosts. In some lungfishes (Neoceratodus), two abdominals fuse to form a single ventral abdominal vein which terminates into sinus venosus. Blood from liver is taken to sinus venosus through a pair of hepatic veins or sinuses.
- Tetrapods :- Embryonic tetrapods also exhibit anterior, posterior and cardinal veins. In adult tetrapods, anterior cardinals persist as the internal jugular veins. The inferior jugular are absent. Common cardinals become the anterior venae cavae or precavals which join sinus venosus (amphibians, reptiles) or directly enter the right auricle of heart (birds, mammals) when a sinus venosus is lacking. In some mammals, (man, cat) left precaval disappears, so that blood of left side enters right precaval through a vessel.
In adult Necturus and larval frog, the postcardinals retain primitive condition, joining caudal vein posteriorly and common cardinal (precaval) anteriorly. In tetrapods, anterior part of each postcardinal disappears but partially present in reptiles, birds and mammals under new names, such as azygos, hemizygos and so forth. Whereas, the posterior part of postcardinal, in continuation with the caudal vein, forms the renal postal vein.
With the suppression of postcardinals, a new vessel, called inferior vena cava or postcaval, develops in tetrapods. It is a large median vessel between the two kidneys originating in the embryonic subcardinal venous plexus. It conveys blood of hind limbs, tail, kidneys, and liver into sinus venosus (amphibians, reptiles) or into right atrium (birds, mammals).
In amphibians, two embryonic ventral or anterior abdominal veins become fused in the adult to form a single median ventral abdominal vein. But it terminates anteriorly into liver and no longer drains the forelimbs. In reptiles, abdominal veins remain paired throughout life and also terminate into liver. They remain connected anteriorly with the hepatic portal system, and posteriorly with the renal portal system (by external iliac). In birds, they are modified into epigastric and coccygeo-mesenteric veins. In mammals, abdominal veins are absent except in spiny anteater (Tachyglossus).
In air-breathing vertebrates, pulmonary veins drain the lungs and enter the left auricle. In lungfishes and amphibians, right and left vessels unite to form a common pulmonary vein opening into right auricle.
2. Renal portal system Blood collected from capillaries in different parts of body is returned directly to the heart through systemic veins and their tributaries. However, in some cases, the returning blood is forced to run through a secondary capillary network in kidneys or liver before being sent to the heart. This is called portal circulation. The vein carrying blood from one set of capillaries to another is termed a portal vein. All the constituents in a portal circulation together form a portal system, named after the organ of body having the secondary capillary’ network. Two portal systems exist in vertebrates : (i) renal portal and (ii) hepatic portal.
Renal portal system is not universally present in all vertebrates. Cyclostomes have no renal portal system. In vertebrates embryos, caudal vein trifurcates anteriorly into a subintestinal and two postcardinal veins, (basic pattern).
In fishes, connection of caudal vein with subintestinal is lost, while the anterior parts of postcardinals are suppressed. As a result, the persistent posterior parts of postcardinals become renal portal veins which pour all blood from tail into kidneys through afferent renal veins. Inside kidneys, they contribute blood to the capillary network surrounding the mesonephric tubules, but never to the glomeruli. Thus renal portal system drains only the tail in fishes.
In amphibians and reptiles, an external iliac vein connects the renal portal and abdominal veins, so that the renal portal system drains the tail as well as the hind limbs. In tailless amphibians or anurans (frog), it drains only hind limbs, while in snakes, having no limbs, it drains only the tail. In Necturus, renal portal vein is directly continuous with postcardinal, as in the basic pattern.
In crocodilians and birds, renal portal system is degenerate and bypasses the kidneys. Only a very little venous blood enters the capillaries in kidneys while most blood from tail and hind limbs passes nonstop through kidneys and leads directly into the postcaval. In mammals, renal portal system is completely absent, except in monotremes, since blood from tail and hind limbs is drained solely by the postcaval.
Significance :- During metabolic activities in the body certain toxic or harmful end products such as urea, uric acid, ammonia, etc, are formed. Renal portal system sends blood from hinder body region directly to the kidneys for the removal of these waste products.
3. Hepatic portal system :- Hepatic portal system is of universal occurrence and essentially similar in all vertebrates. In the embryo, the first venous channels to form are a pair of vitelline veins (basic pattern) arising from the yolk sac or midgut to enter the sinus venosus of heart. Caudal vein from tail is continued forward beneath the digestive tract as a subintestinal vein, which usually joins the left vitelline vein. As liver develops, vitelline veins unite together forming a single hepatic portal vein in lesser omentum. The subintestinal vein also loses connection with caudal vein to become a part of hepatic portal system. It drains different parts of digestive tract (yolk sac, stomach, intestine, etc.), its various derivatives (pancreas, gall bladder, rectal gland, etc.) and spleen, and passes it on to the sinusoids in liver. In adult sharks, vitelline veins remain paired and form hepatic sinuses.
Significance :- Intestinal capillaries absorb several kinds of dissolved food material, except fats, from alimentary canal. These are carried in blood through hepatic portal vein to liver to be temporarily stored up until required. Carbohydrates and sugars are converted into glycogen to be stored in liver cells. If all the sugar remains in circulation, the concentration of blood sugar may go up leading to diabetes. Stored sugar from liver cells is released whenever sugar content of blood drops below normal, which may otherwise lead to unconsciousness. Liver also converts proteins and amino-acids into urea and renders carbolic acid, cresol, indol, ammonia, etc., harmless.
See Other Posts Also
- Circulatory System in Vertebrates
- Digestive System in Vertebrates
- Integument or Skin and its Derivatives in Vertebrates
- Placenta in Mammals (Placentation)
Also Follow Us on
Discover more from Zoologyverse
Subscribe to get the latest posts sent to your email.